Consciousness, the ability to observe, includes an awareness that observation occurs. It describes not what we observe but that we observe, in a subjective experience that others confirm. In contrast, there is no evidence that a camera experiences the photos it takes. It is just a set of atoms, with no subject to see what it makes. If our brain is a biological camera, it should be the same, so is the observation experience imaginary?
I think I observe, but am I imagining an event that doesn’t occur? In scientific terms, either observation is a real event, where a subject observes an object, or it isn’t. Whether consciousness is imaginary or not then depends on whether the act of observation occurs.
Current neuroscience doesn’t agree that the brain makes one observation because it has no physical center to do that. For example, reading this sentence involves brain areas of vision, language and thought that have no central control. Without a control center, the text, grammar, and meaning of a sentence can’t form one observation, but it happens. I observe the appearance, syntax, and semantics of a sentence in parallel, not sequentially, and can focus attention on any aspect at will. The brain is like a city with an aerial observer who sees the whole yet can jump to any location detail, regardless of the road links! A brain with no control center shouldn’t be able to do this.
And to conclude that we don’t observe at all is also a problem, because what then is science based on? How do we know anything at all if observation isn’t real? If the act of observing is imaginary, then so are all our facts. Rejecting observation denies both our experience of it and science, including quantum and relativity theories, so most scientists accept that we do observe, but if so, to what event does it refer (Note 1)?
To observe as we do is to experience the state of something else, not just process information as cameras do. We don’t just register red light frequencies; we see redness itself. A subject observes and knows that it does so, but physical things can’t both act on others and experience them, so that they observe is inherently illogical.
Physical events can’t explain observation but quantum theory can, as it attributes it to quantum collapse. Observation is said to occur when quantum waves collapse in a physical event that restarts them at the same point, to become one. Being the same entity lets them experience each other, and they retain that knowledge when they disentangle, and that is observation. Note that what observes isn’t the waves that spread, but the entities that generate and restart them (6.3.16).
Physical realism makes observation imaginary but quantum realism makes it real. It lets our brains entangle millions of nerves into an entity that can observe and choose for the whole, with obvious evolutionary benefits. And if observation is a real event, mystics meditating on it are studying reality, so their honest reports:
“…should claim from us the same attention we give to other explorers of countries in which we are not competent to adventure ourselves; for mystics are the pioneers of the spiritual world…” (Underhill, 1961) p4.
If the spiritual world is an experience of quantum reality (7.1.1), statements about it should satisfy quantum theory. For example, Hindu scriptures say that the divinity within has no parts, nor is it tainted by physical events:
“In the supreme bright sheath is Brahman, free from taints and without parts.” (Mundaka Upanishad, II.ii.9).
If Brahman (the divinity within) is a quantum entanglement made by the brain, then it indeed has no parts because quantum theory describes entanglement as a genuine unification, with no parts at all. And if this observer exists outside physical space, it is beyond all physical taints. This scripture is then valid in the sense that it doesn’t contradict quantum theory, and it is reliable if other reports repeat it. In contrast, claims that divinity enables powers like flying aren’t valid because no evidence suggests that quantum entities can nullify gravity. That we observe by quantum means isn’t a carte blanche for any claim because quantum reality must follow quantum laws.
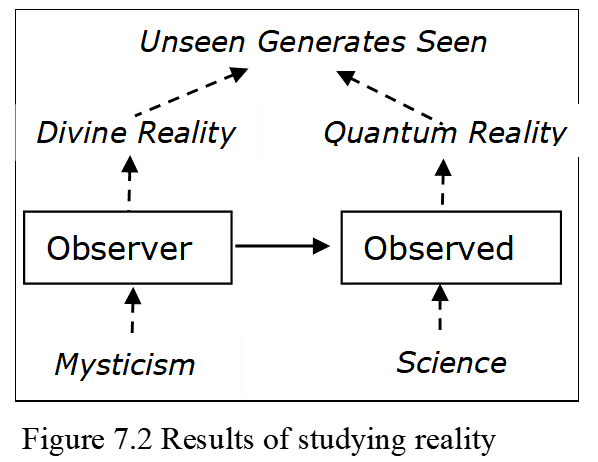
Quantum realism concludes that quantum reality causes matter and the ability to observe it, and scriptures say the same of divinity. Current science calls both imaginary but the evidence disagrees, as quantum events explain matter and observation better than physical events (Chapters 1-6). In our observer-observed reality, the study of either suggests that the unseen causes the seen (Figure 7.2). Mysticism and science both tend to conclude that the unmanifest became manifest by physical events. And if observation is an event that occurs, albeit in the quantum realm, there must be beings that observe. Regardless of what is observed, observation needs a being to do so.
In this view, consciousness is the ability to observe and being is that which observes. There is being and manifestation, one real and the other not. Being, the entity that observes and chooses, is real in the sense that it exists both before and after physical events. In contrast, manifestation, the appearance to us of things, is unreal in the sense that they don’t exist before or after physical events. Beings then generate quantum waves that interact in physical events that let them observe each other.
That being isn’t physical supports its survival but that it needs brain waves that end at death doesn’t. Either way, consciousness is fundamental to nature because every physical event is a mutual observation. If everything observes on some scale, consciousness is all around us, just as light is. The eyes don’t create light, they just focus it, and likewise the brain doesn’t create consciousness, it just concentrates it. The brain is like a lens that condenses light, where:
“Allah (God) is the light of the heavens and the earth.” (Koran, 4.35).
If our eyes fail, there isn’t less light in the universe, and equally if our brains fail, the consciousness of the universe isn’t reduced. Being was given to us, just as light was, so the question isn’t whether the primal reality that made our universe billions of years ago will survive our death, but whether what our brain made of it can. The next sections examine this and other questions raised by scriptures and their mystical traditions, given that consciousness is part of the natural world, and under the right circumstances, it can coalesce into beings like us.
Note 1. Facing this problem, some materialists suggest that observation is an unknown property of matter (Strawson, 2008), but even this can’t explain how the observations of atoms became human observations.